
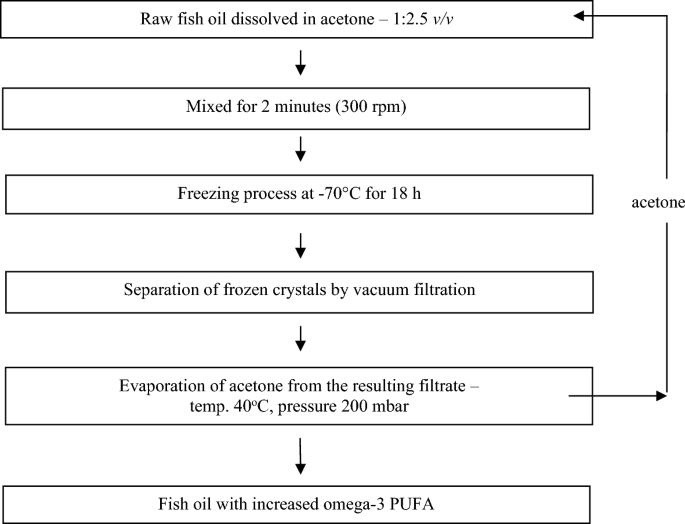
Composition and fatty acid profile of milk from cows fed diets supplemented with raw and n-3 PUFA-enriched fish oil
Omega-3 LC-PUFAs are chemically unstable and highly susceptible to oxidation due to their high number of double bonds and their geometrical position within the FA chain. The oxidation process results in an unpleasant taste and odour, as well as the formation of toxic substances and unfavourable changes in the configuration of the FAs from cis to trans35. To reduce the adverse oxidative changes in mackerel oil enriched in n-3 PUFAs, Patkowska-Sokoła et al.25 used α-tocopherol as an antioxidant and found a slight decrease in EPA and DHA levels and a slight increase in peroxide and anisidine levels after 4 weeks. In order to limit the adverse oxidative changes in our study, an analogous amount of alpha-tocopherol as antioxidant and stored both preparations (LTC-FO, FO) under identical conditions were used. In order to facilitate the use of LCT-FO and FO in cattle feeding, both formulations were sprayed on the carrier Humokarbowit, which is characterised by high sorptive capacity and antioxidant properties and is suitable for livestock due to its biostimulatory and prophylactic properties14,36.

Due to the lack of reports in the available literature on the use of LC-PUFA-enriched fish oil in ruminant diets, the discussion section compares the results obtained in the present study with studies on dietary supplementation with different amounts of fish oil.
In our experiment, the addition of LTC-FO resulted in a lower intake of DM compared to FO. A decrease in DMI with increasing levels of fish oil in the ration of cows was also observed by other authors. In the study by Donovan et al.16 at 1, 2 and 3% fish oil addition, DMI was 29.0, 23.5 and 20.4 kg/d, respectively. In the experiment of Keady et al.13 in the groups receiving 150, 300 and 450 g/d of fish oil, silage dry matter intake was 10.2, 9.2 and 7.9 kg/d, respectively. Also Kairenius et al.8 reported that after rumen administration of 75 and 300 g/d of fish oil, intake of silage and total DM was 10.8 and 18.9 kg/d, and 8.25 and 16 kg/d, respectively.
In our study, supplementation of dairy cows’ diets with LTC-FO compared to FO had no effect on milk production, lactose content and yield and protein content, only a decrease in fat content and yield and protein yield in milk was found (Table 3). The results of other authors’ studies on the effects of different doses of fish oil in the feeding of cows on milk yield, protein and lactose content were inconclusive8,13,16. In most studies, fish oil supplementation, especially in the unprotected form, typically reduces milk fat synthesis in lactating cows. Donovan et al.13 showed that fat content and yield decreased linearly with increasing fish oil dose from 1 to 3%. In the study by Kairenius et al.8, fish oil supplementation at levels of 75, 150 and 300 g/day reduced milk fat content (by 5.6%, 19.9% and 30.1%, respectively) and fat yield (by 5.2%, 15.1% and 40.6%, respectively). Rego et al.37 observed decreases in milk fat content of 4.9 and 11.4% and fat yield of 140 and 340 g/day, respectively, at 160 and 320 g fish oil supplementation. Diet-induced milk fat depression (MFD) is characterised by a significant reduction in fat content due to changes in rumen metabolic pathways, with no change in yield or other milk constituents38. For diets containing marine supplements, the biohydrogenation (BH) theory has been proposed, which attributes the reduction in milk fat to the formation of specific FA intermediates, which after absorption in the duodenum and transfer to the mammary gland, inhibit milk fat synthesis38,39,40. Changes in the FA profile associated with FO-induced MFD clearly indicate that the decrease in milk fat content to inhibition of de novo FA synthesis rather than preformed FA uptake in the mammary gland41,42. A typical phenomenon associated with the use of fish oil supplements in cattle feed is an increase in trans-C18:1 isomers in milk fat38,39,43. Numerous studies have shown that FO-induced MFD is also associated with a reduced proportion of 18:1 cis-9 in milk fat44,45. In our study, as a result of supplementation with LTC-FO compared with FO, the proportion of C18:1 trans-10 and trans-11 in milk fat increased and the proportion of C18:1 cis-9 decreased, which may explain the lower fat content in the LTC-FO group.
Milk fat synthesis depends on two general sources of FAs. The short- (SCFA) and medium-chain (MCFA) FAs (C4–C14) and half of the C16 FAs are synthesised de novo in the mammary gland, while FAs > 18-carbon are transferred from preformed blood triglycerides46. As a result of feeding fish oil to ruminants, changes in rumen BH pathways occur towards reduced content of milk fat SFA and increased LC-PUFA8,47. Fish oil supplementation decreased the proportion of 4–16-carbon FAs in milk38,39, which may be explained by the inhibitory effects of increased long-chain FAs availability on genes involved in lipid metabolism in the mammary gland48. Previous studies have shown that fish oil results in a dose-dependent decrease in the proportion of FAs synthesised de novo in lactating cows8,13,37.
In our research, the levels of butyric acid (C4:0), caproic acid (C6:0), caprylic acid (C8:0) and capric acid (C10:0) in the SCFA group decreased as a result of supplementing cows’ diets with LCT-FO vs. FO (Table 4). In the experiment by Kairenius et al.8 the levels of C4:0, C6:0, C8:0 and C10:0 were lower when fish oil was added at 150 and 300 g/d than at 75 g/d. In the studies by Keady et al.16, Donovan et al.13 and Rego et al.37, a greater decrease in C6:0, C8:0 and C10:0 was observed with increasing fish oil dose. Conversely, in our study in the MCFA group, the lauric acid (C12:0) and palmitic acid (C16:0) content decreased and the palmitoleic acid (C16:1 cis-9) content increased (Table 4), which is confirmed by the results of the other studies as the level of FO in the dose increases8,37.
Under normal conditions, stearic acid (C18:0) is the major FA leaving the rumen during the BH process49. Supplementation of ruminant diets with marine lipids inhibits the final step of BH to C18:0 in the rumen50, which reduces its uptake by the mammary gland and consequently reduces the synthesis of oleic acid (18:1 cis-9) by delta(9)-desaturase51. In addition, the use of marine lipids results in an increase in trans-18:1 isomers in milk fat, which have a higher melting point than cis-C18:1 isomers8,21. As a result of LTC-FO vs. FO supplementation, the content of C18:1 cis-9 in milk fat decreased and the content of C18:1 trans-10 and trans-11 increased (Table 4). In the experiment by Shingfield et al.52, fish oil increased the content of trans FA in milk, probably due to incomplete BH of unsaturated 16- to 22-carbon FA in the rumen. In vitro studies have shown that both EPA and DHA inhibit the reduction of 18-carbon unsaturated FA to 18:0, resulting in the accumulation of trans-18:1 intermediates53,54. Loor et al.21 and Shingfield et al.55 reported that most of the increase in total milk trans FA content due to fish oil supplementation was associated with a specific enrichment of trans-18:1 isomers (Δ8–12). The linear increase in the content of C18:1 isomers of the configurations trans-10 and trans-11 (TVA) with increasing fish oil dose was observed in the study by Kairenius et al.8, TVA in the research by Donovan et al.13 and total trans-C18:1 isomers in the experiment by Keady et al.16.
Marine lipids with high content of 20- and 22-carbon PUFA have often been used to inhibit the saturation of 18:1 trans-11 with the ultimate aim of increasing C18:2 cis-9,trans-11 in milk8,56. In our experiment a higher level of the CLA isomer cis-9,trans-11 (RA, rumenic acid) was observed in the LTC-FO group compared to the FO group (Table 4). An increase in RA content was also observed in the studies by Kairenius et al.8 (1.03 vs. 2.15 g/100 g total FA, 75 vs. 150 g/d fish oil), and Donovan et al.13 (1.58 vs. 2.23 g/100 g total FA, 1 vs. 2% fish oil).
Supplementation with LTC-FO resulted in an increase in the content of α-linoleic acid (ALA, C18:3 all-cis-9,12,15; n − 3) and γ-linoleic acid (GLA, C18:3 all-cis-6,9,12; n − 6) compared to FO supplementation (Table 4). This is consistent with previous studies, such as Keady et al.16, where GLA content increased with higher levels of fish oil intake (150–300 g/d). Kairenius et al.8 also found an increase in ALA content with the highest dose of fish oil. In contrast, Donovan et al.13 found that the higher level of fish oil in cows’ feed had no effect on the content of ALA, and caused a decreased in GLA.
The supplementation of LTC-FO vs. FO resulted in an increase in the content of most long-chain PUFAs (≥ 20-carbon) found in milk fat. These include eicosanoic acid (C20:1 cis-11), eicosadienoic acid (EDA, C20:2 cis-11,cis-14; n − 6), dihomo-γ-linolenic acid (DGLA, C20:3 all-cis-8,11,14; n − 6), and arachidonic acid (AA, C20:4 all-cis-5,8,11,14; n − 6) (Table 4). Previous research has shown that increasing the amount of fish oil in the diet leads to an increase in the milk fat content of C20:1, C20:2, C20:3 and C20:48, C20:1 and C20:413, and C20:1, C20:2 and C20:416.
Eicosapentaenoic acid (EPA, C20:5 all-cis-5,8,11,14,17; n − 3) and docosahexaenoic acid (DHA, C22:6 all-cis-4,7,10,13,16,19; n − 3) are typically present in very low amounts in milk due to their absence or minimal levels in traditional dairy cow diets13,20. The transfer of dietary EPA and DHA to milk fat in lactating cows is very low (< 4%)19, reflecting extensive BH by rumen bacteria21,52. The low transfer may also be a result of the partitioning of EPA and DHA into plasma lipid fractions that are less available to the mammary gland, such as triglycerides and non-esterified fatty acids57. Additionally, after absorption, EPA and DHA are incorporated into cholesterol esters and phospholipids that have a low affinity for lipoprotein lipase in the mammary endothelium58. The BH process activity decreased when fish oil was added to the animals’ feed rations. This is likely due to the toxic effects of PUFAs on certain rumen bacteria20,40,52. Previous studies have shown that supplementing with fish oil increases the content of milk C20:5, C22:5, and C22:6 in a dose-dependent manner8,13,16. The effective transfer of these fatty acids is greater when they are delivered in a way that bypasses rumen metabolism56. In our experiment, adding LTC-FO instead of FO resulted in an increase in milk fat EPA, DHA, and docosapentaenoic acid (DPA, C22:5 all-cis-7,10,13,16,19; n − 3) (Table 4). Rego et al.37 conducted an experiment where they found that the content of EPA in milk fat increased by 2.6- and 4.7-fold (from 0.007 to 0.18 and 0.33 g/100 g FA, respectively), and DHA by 2.8- and 7.2-fold (from 0.06 to 0.17 and 0.43 g/100 g FA, respectively), for treatments of 160 and 320 g/d of fish oil. Similarly, Keady et al.16 demonstrated that EPA content was significantly higher with fish oil supplementation of 450 g/d compared to 150 or 300 g/d. Kairenius et al.8 found that milk fat from cows supplemented with 300 g/d fish oil had significantly higher contents of EPA, DHA, and DPA than that supplemented with 75 or 150 g/d.
When analyzing changes in the content of FA groups with varying degrees of saturation and carbon chain length, in the LTC-FO fed cows a decrease in the proportion of SFAs in milk fat was observed, including short-chain SCFAs, and an increase in the proportion of UFAs, PUFAs, and LC-PUFAs (Table 5). This finding is consistent with other reports. Donovan et al.13 reported a decrease in the proportion of SFAs and SCFAs, and an increase in the proportion of UFAs in cow’s milk as a result of increased addition of fish oil. Similarly, Kairenius et al.8 observed a linear decrease in the proportion of SFAs and an increase in PUFAs in milk fat with an increase in the level of fish oil in cows’ diet. Rego et al.37 showed that increasing dietary fish oil intake led to a decrease in total SFAs and the proportion of short-chain fatty acids (C4:0 to C12:0), while the proportion of medium-chain fatty acids (C14:0 to C16:1) remained unchanged.
As a result of LTC-FO compared to FO supplementation, total CLA, trans-18:1 isomers, n-3 FAs and EPA + DHA in milk fat increased (Table 5). As in our study, Kairenius et al.8 found an increase in total CLA and trans isomers in milk fat and Donovan et al.13 found an increase in total n − 3 FAs with an increase in fish oil supplementation in cows’ diets. Conversely, in the study by Rego et al.37, an increase in fish oil supplementation in cows’ diets was associated with an increase in EPA + DHA in milk fat.
Functional food is the fastest growing sector of the global food market59. Cow’s milk is a strategic commodity and a staple food. Modification of the FA profile by supplementation with fish oil or its combination with plant oils is a good dietary strategy to reduce the intake of SFAs in favour of healthier FAs (e.g. n − 3 LC-PUFA, CLA)8,14,60,61. Studies conducted in patients with myocardial infarction, metabolic syndrome and vascular disease describe benefits and suggest that milk enriched with n − 3 LC-PUFA (EPA and DHA) may be useful as a dietary supplement to control risk factors62.
In 2022 the EU average of milk consumption was about 53.4 kg per person, which is about 140 g/d63. Consuming an equivalent amount of milk from LTC-FO-supplemented cows can provide a person with 388 mg EPA and 360 mg DHA, while from FO-supplemented cows 297 mg and 294 mg, respectively. Recommendations for daily EPA + DHA intake depend, among other factors, on age (children, adult, elderly), gender (men, women), health status (for cardiovascular health), physiological status in women (pregnant, lactating), and range from 250 to 670 mg/d. For healthy adults (19–57 years), the minimum intake should be 500 mg/d64,65. This amount of EPA + DHA in a person’s diet can provide an intake of approximately 95 g/d of milk from cows supplemented with LTC-FO.
Source link









Share this article: